A fisiologia do endurance revela, em sua própria lógica biológica, por que intervenções que ampliam a entrega e o uso de oxigênio alteram de maneira tão profunda o rendimento humano. A performance em esforços prolongados depende de um eixo integrado que começa no coração, passa pelo sangue e culmina no músculo esquelético, onde o oxigênio é finalmente convertido em energia mecânica. Entre todos esses componentes, o mais maleável é o sistema hematológico, especialmente a capacidade de transportar oxigênio por meio da hemoglobina. Essa vulnerabilidade fisiológica — que não é uma falha, mas uma característica evolutiva — explica por que o doping sanguíneo permanece tão eficaz e tão difícil de combater. Organismos aeróbios, desde peixes primitivos até mamíferos, evoluíram estratégias para maximizar o uso de oxigênio, mas nunca desenvolveram defesas contra aumentos artificiais e abruptos na massa de glóbulos vermelhos. É justamente nesse “espaço evolutivo” que o doping atua. Ainda nos primeiros vertebrados, as hemácias eram enormes e lentas; sua redução progressiva ao longo da evolução tornou o transporte de oxigênio mais rápido e eficiente, mas deixou o sistema vulnerável a manipulações artificiais que o organismo nunca precisou “prever”, criando uma brecha que hoje é explorada com precisão bioquímica.
A base fisiológica do rendimento aeróbio pode ser entendida pela equação clássica do transporte de oxigênio: o débito cardíaco máximo tende a atingir limites rígidos mesmo em atletas treinados; a extração muscular já opera próxima ao máximo durante esforços intensos; e a única variável realmente expansível é o conteúdo arterial de oxigênio, determinado pela hemoglobina. Quando a massa total de hemoglobina aumenta — seja por transfusão, estímulo farmacológico à eritropoiese ou manipulação genética — o corpo passa a entregar mais oxigênio aos músculos com menor estresse para cada batimento cardíaco. Essa simples modificação hematológica produz um encadeamento fisiológico poderoso: maior VO₂máx, menor produção de lactato para a mesma velocidade, maior economia mecânica, maior tempo até a exaustão e maior preservação de glicogênio. Do ponto de vista evolutivo, é como se o atleta adquirisse temporariamente um fenótipo adaptado à altitude, porém sem qualquer custo biológico real de adaptação. A separação entre circulação sistêmica e pulmonar, que surgiu tardiamente na evolução, permitiu a formação de capilares cada vez mais estreitos e densos; isso aumentou drasticamente a eficiência da troca gasosa, mas também criou um cenário no qual qualquer aumento artificial da hemoglobina amplifica esse sistema muito além do seu regime natural, rompendo limites que a seleção natural jamais teve motivo para impor.
O doping sanguíneo clássico — especialmente a autotransfusão — explora essa fragilidade do sistema. Ao remover e reinfundir o próprio sangue, o atleta aumenta diretamente a massa de hemácias circulantes. Mesmo volumes discretos, de cerca de uma xícara de hemácias concentradas, elevam de forma perceptível a capacidade aeróbia. O organismo não dispõe de mecanismos eficientes para detectar “excesso de oxigênio”, porque, ao longo da evolução, nunca foi ameaçado por isso. Assim, uma autotransfusão produz um estado hematológico supra-fisiológico, impossível de ser alcançado por treinamento ou genética natural. Pequenas reinfusões, chamadas microtransfusões, são particularmente problemáticas porque provocam aumentos contínuos e quase imperceptíveis de hemoglobina, evitando alterações abruptas que poderiam ser detectadas. O músculo passa a receber mais oxigênio por unidade de fluxo sanguíneo, o que reduz o recrutamento de vias anaeróbias e adia o ponto em que o lactato se acumula. Desde os peixes primitivos, cuja pressão arterial era baixíssima, a eficiência do transporte sanguíneo aumentou com a miniaturização das hemácias; nos mamíferos modernos, essa delicadeza estrutural permite que pequenas manipulações aumentem de forma desproporcional a entrega de oxigênio, fazendo com que intervenções mínimas gerem ganhos fisiológicos descomunais.

Imagem: Lance Armstrong won the Tour de France seven times in a row, but the results have now been voided. https://www.newscientist.com/article/2139340-blood-doping-were-armstrong-and-russia-wasting-their-time/
A eritropoietina sintética opera em outro ponto da cadeia: ela estimula a medula óssea a produzir mais glóbulos vermelhos, ativando vias intracelulares que inibem a morte dos precursores e aceleram sua proliferação. Essa sequência envolve ativação de receptores, cascatas de fosforilação, aumento de proteínas antiapoptóticas e expansão dos reticulócitos em poucos dias. O resultado final é um aumento progressivo e contínuo da capacidade de transporte de oxigênio. Microdoses de EPO têm se tornado comuns justamente porque modulam discretamente essa via: ao administrar pequenas quantidades repetidas, o atleta alimenta constantemente a zona mais sensível dos precursores eritroides, ampliando a produção sem gerar variações marcantes nos biomarcadores monitorados. Como efeito paralelo, a queda nos níveis de hepcidina aumenta a disponibilidade de ferro, acelerando ainda mais a maturação dos glóbulos vermelhos. É um ciclo fisiológico otimizado artificialmente, mas que o organismo aceita como se fosse um estado natural de maior necessidade de oxigênio. A medula óssea, que substituiu a produção de hemácias no fígado e baço dos vertebrados mais antigos, nunca foi selecionada para responder a estímulos farmacológicos — apenas à hipóxia real; por isso reage “obedientemente” à EPO sintética como se o corpo estivesse vivendo em altitude extrema, reinterpretando sinais artificiais como pressões adaptativas legítimas.

Imagem: https://www.outsideonline.com/health/training-performance/biological-passport-doping-study/
O doping genético representa uma fronteira ainda mais profunda porque altera a lógica molecular do próprio organismo. Estabilizar moléculas reguladoras sensíveis ao oxigênio ou introduzir genes que elevam de forma contínua a produção endógena de EPO cria um ambiente celular onde a medula óssea interpreta uma hipóxia inexistente. Isso altera o fenótipo hematológico por meses, sem picos detectáveis e sem as assinaturas bioquímicas deixadas por substâncias externas. Além disso, manipulações gênicas podem atuar não apenas no sangue, mas nos tecidos-alvo da performance: músculo esquelético, mitocôndrias, fibras oxidativas, sistemas de tamponamento e até vias de angiogênese. Ao modificar diretamente a arquitetura metabólica e estrutural do músculo, o doping genético não apenas melhora o desempenho, mas reconstrói o atleta como um organismo biologicamente distinto. Enquanto a evolução natural levou milhões de anos para aperfeiçoar gradualmente capilares, mitocôndrias e fibras musculares oxidativas, o doping genético recria esses saltos em meses — uma velocidade sem precedentes na história evolutiva dos vertebrados — produzindo adaptações “instantâneas” que jamais ocorreriam naturalmente.

Imagem: Kaarlo Maaninka (208), the subject of the first known blood doping case, in the 1980 Summer Olympics 5,000 m race. https://en.wikipedia.org/wiki/Blood_doping
A introdução do monóxido de carbono como ferramenta ergogênica recente expõe outra brecha fisiológica herdada dos vertebrados: a extrema sensibilidade da eritropoiese a reduções mínimas na oferta de oxigênio renal. A inalação controlada de CO, mesmo em microdoses de 1 mL/kg, aumenta rapidamente a fração de carboxi-hemoglobina (HbCO), deslocando o oxigênio das hemácias e criando um estado de “hipóxia funcional” que o organismo interpreta como ameaça legítima. Esse pequeno desvio na saturação é suficiente para reduzir a oxigenação do córtex renal — região evolutivamente selecionada para detectar quedas ínfimas de O₂ — desencadeando um pico de eritropoietina entre 2 e 4 horas após a exposição, como já foi demonstrado experimentalmente com a elevação de 42% de EPO mesmo após uma única inalação.
O CO, ao ocupar transitoriamente os sítios da hemoglobina, gera um sinal bioquímico que a evolução nunca precisou calibrar contra manipulações externas; por isso, a medula responde de forma amplificada, acelerando a maturação eritroide, aumentando reticulócitos e expandindo o volume total de hemoglobina em poucas semanas. Mais ainda: modelos recentes mostram que exposições intermitentes podem elevar a tHb em magnitude comparável ao treinamento em altitude, atingindo aumentos de até 4,8% em três semanas, confirmando que o eixo O₂-rim-medula permanece vulnerável a perturbações artificiais de pequena escala.
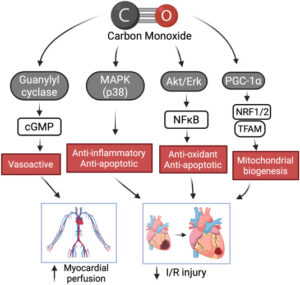
Imagem: Chu LM, Shaefi S, Byrne JD, Alves de Souza RW, Otterbein LE. Carbon monoxide and a change of heart. Redox Biol. 2021;48:102183.
Essa intervenção revela um paradoxo evolutivo ainda mais profundo: ao reduzir a capacidade imediata de transporte de oxigênio pelo aumento da HbCO, o CO desencadeia adaptações que ampliam de maneira duradoura exatamente o mesmo sistema que inicialmente compromete. A capacidade total de hemoglobina aumenta, o volume plasmático se expande e, como observado em atletas treinados, o VO₂máx eleva-se de forma mensurável após semanas de microexposição repetida.
Paralelamente, estudos recentes descrevem que o CO altera a cinética da curva de dissociação da hemoglobina, deslocando-a para a esquerda, o que impacta a dinâmica de liberação de O₂ nos tecidos — um ajuste transitório, mas suficiente para amplificar os sinais renais de “déficit” e intensificar a eritropoiese. Ao mesmo tempo, surgem efeitos secundários no músculo, incluindo maior densidade mitocondrial, aumento de mioglobina e potenciação da angiogênese, reforçando um perfil oxidativo semelhante ao de espécies adaptadas a ambientes hipóxicos crônico. Assim, a inalação de CO não apenas manipula o sangue; ela recria artificialmente um cenário evolutivo que acelera e condensa pressões seletivas que levaram milhões de anos para moldar o aparato aeróbio dos vertebrados, inserindo mais uma fissura explorável na lógica biológica do transporte de oxigênio.

Enquanto o doping sanguíneo altera principalmente o transporte de oxigênio, os esteroides anabolizantes atuam sobre o outro grande pilar do endurance: a capacidade de sustentar volumes extremos de treinamento e de recuperar-se rapidamente. Embora classicamente associados à força, seus efeitos mais relevantes para provas longas aparecem na biologia da reparação muscular. Ao aumentar a síntese proteica e reduzir o catabolismo, eles diminuem o dano estrutural após sessões intensas. Isso permite acumular mais horas semanais de treino — e no endurance, volume é talvez o maior determinante da adaptação crônica. Além disso, alguns esteroides aumentam discretamente a eritropoiese, criando sinergia com métodos que expandem a capacidade aeróbia. A combinação entre elevação da hemoglobina e maior resiliência muscular dá ao atleta um perfil fisiológico duplamente ampliado: ele pode produzir mais energia e, simultaneamente, tolerar mais treinamento, gerando um ciclo de aprimoramento artificial que não tem paralelo na biologia natural. Fibras musculares altamente oxidativas surgiram tarde na evolução dos vertebrados; reforçá-las artificialmente com hormônios acelera e amplifica processos que levaram milhões de anos para se estabelecer, comprimindo eras evolutivas em ciclos de semanas.
O panorama epidemiológico reforça essa realidade fisiológica. A prevalência real de doping em provas de endurance é muito maior do que sugerem estatísticas oficiais, especialmente em modalidades de longa duração. A disparidade entre o índice real e o número de casos detectados evidencia que as estratégias atuais de controle capturam apenas intervenções grosseiras. Microdosagens de EPO, autotransfusões fracionadas, manipulações de volume plasmático e técnicas emergentes dificultam profundamente a detecção. O passaporte biológico melhora a vigilância, mas permanece vulnerável porque se baseia em marcadores que variam naturalmente entre indivíduos e são sensíveis a fatores não relacionados ao doping, como altitude, hidratação, calor e variações sazonais. Como as hemácias apresentam ciclos naturais de variação herdados de nossos ancestrais vertebrados — influenciados por temperatura, disponibilidade de oxigênio e até ritmos circadianos — essas flutuações mascaram interferências artificiais, criando zonas cinzentas que desafiam a sensibilidade dos modelos estatísticos mais modernos.
A convergência de todos esses métodos — sangue, hormônios, genes e anabolizantes — cria um ecossistema fisiológico inteiramente novo. O doping moderno se tornou uma estratégia sistêmica, não um ato isolado. O atleta que manipula sua eritropoiese tende a usar esteroides para treinar mais, moduladores metabólicos para suportar intensidades extremas e táticas genéticas para alterar a própria lógica de regulação do oxigênio. Do ponto de vista biológico, o resultado é a criação de um organismo desenhado para sustentar níveis de estresse fisiológico incompatíveis com os limites naturais da espécie humana. Nenhum vertebrado, ao longo de 500 milhões de anos, desenvolveu simultaneamente máxima capacidade aeróbia, máxima densidade capilar e máxima regeneração muscular — esses atributos nunca coexistiram naturalmente em um só organismo; e, ainda assim, o doping força sua convergência em um único corpo, algo sem precedentes na história natural.
No fim, a eficácia extraordinária do doping em endurance não deriva de truques farmacológicos sofisticados, mas da própria estrutura da biologia humana. Nossa fisiologia aeróbia evoluiu para otimizar o uso de oxigênio dentro de limites estreitos, não para lidar com aumentos artificiais de hemoglobina, manipulação da medula óssea ou remodelação genética. O doping explora justamente esses pontos frágeis — e é por isso que seus efeitos são tão grandes e tão difíceis de neutralizar. Ao ampliar de forma artificial o eixo que sustenta a capacidade de transporte de oxigênio e ao reforçar a resiliência muscular ao treinamento, ele reorganiza o corpo humano em um sentido que ultrapassa o treinamento, criando um fenótipo de performance biologicamente impossível de atingir por vias naturais. Como mostram os registros evolutivos — hemácias menores, capilares mais densos, pressões arteriais mais altas, corações mais seccionados — todos esses avanços ocorreram lentamente; o doping rompe essa cadência, criando um “supervertebrado” que nunca existiu na história natural, reunindo num mesmo organismo capacidades que a evolução sempre manteve separadas.
Apesar de todo esse entendimento fisiológico e evolutivo ajudar a explicar por que o doping é tão eficaz, é justamente essa eficácia que reforça a necessidade de sua proibição. As regras esportivas existem para garantir que o desempenho decorra de adaptação biológica legítima, treinamento consistente e talento, e não de intervenções que ampliam artificialmente capacidades que extrapolam o escopo natural da espécie humana. Além disso, mesmo quando administradas sob supervisão médica, essas práticas podem comprometer a segurança do atleta ao alterar sistemas regulatórios finos — hematológicos, cardiovasculares e metabólicos — que evoluíram para operar dentro de margens estreitas. Por isso, condena-se o uso do doping não por um julgamento moral, mas porque ele viola a equidade competitiva, coloca a saúde em risco e rompe o espírito fundamental do esporte: a comparação justa de performances humanas obtidas dentro dos limites naturais e das normas que as regem.
Referencias:
Breenfeldt Andersen A, Nordsborg NB, Bonne TC, Bejder J. Contemporary blood doping-Performance, mechanism, and detection. Scand J Med Sci Sports. 2024;34(1):e14243.
Faiss R, Saugy J, Zollinger A, Robinson N, Schuetz F, Saugy M, Garnier PY. Prevalence Estimate of Blood Doping in Elite Track and Field Athletes During Two Major International Events. Front Physiol. 2020 Feb 25;11:160.
Heuberger JAAC, van Eenoo P, Rotmans JI, Gal P, Stuurman FE, Post TE, Daniels JMA, Ram H, de Hon O, Burggraaf J, Cohen AF. Sensitivity and specificity of detection methods for erythropoietin doping in cyclists. Drug Test Anal. 2019 Sep;11(9):1290-1301.
Jones M, Tunstall Pedoe DS. Blood doping–a literature review. Br J Sports Med. 1989 Jun;23(2):84-8. doi: 10.1136/bjsm.23.2.84.
Marchand A, Ericsson M. Homologous blood transfusion and doping: Where are we now? Drug Test Anal. 2024 Dec;16(12):1479-1486.
Robach P, Trebes G, Buisson C, et al. Prevalence of Drug Use in Ultraendurance Athletes. Med Sci Sports Exerc. 2024;56(5):828-838.
Wang J, Ji Y, Zhou L, Xiang Y, Heinonen I, Zhang P. A New Method to Improve Running Economy and Maximal Aerobic Power in Athletes: Endurance Training With Periodic Carbon Monoxide Inhalation. Front Physiol. 2019 Jun 6;10:701.
Webb KL, González-Alonso J. Heat and carbon monoxide exposure: Is two better than one? Exp Physiol. 2024 Oct;109(10):1627-1628.