A fisiologia do exercício consolidou-se como um campo singular na história das ciências biológicas aplicadas. Ela emergiu da confluência entre o empirismo dos treinadores e a sistematização experimental da ciência moderna, uma autêntica integração entre teoria e prática. Desde os estudos pioneiros de Jöns Jacob Berzelius (1779-1848), que em 1807 identificou lactato em músculos de veados abatidos, sugerindo aumento de sua concentração durante o esforço físico, até as contribuições de August Krogh (1874-1949) e Archibald Hill (1886-1977), o entendimento do metabolismo energético e de suas respostas ao exercício tem evoluído de maneira decisiva. Esse percurso histórico permitiu compreender como o corpo humano regula a produção, o transporte e a utilização de lactato, tornando essa molécula um dos principais marcadores fisiológicos do desempenho atlético.
Embora o conceito de consumo máximo de oxigênio (V̇O₂max) tenha representado um marco na mensuração da capacidade cardiorrespiratória, suas limitações práticas tornaram-se evidentes. A exigência de esforço até a exaustão, o alto custo operacional e a relativa insensibilidade para prever o desempenho em provas de resistência estimularam a busca por indicadores submáximos mais específicos e aplicáveis à realidade do treinamento. Foi nesse contexto que os estudos sobre os limiares de lactato e ventilatórios se tornaram centrais para a compreensão das transições metabólicas durante o exercício.
Na década de 1950, Wildor Hollmann (1925-2021) introduziu o conceito de “ponto de eficiência ótima da ventilação”, um precursor das modernas avaliações de limiares metabólicos. Embora seu trabalho tenha inicialmente permanecido restrito ao meio acadêmico de língua alemã, abriu caminho para o reconhecimento das transições entre metabolismo aeróbico e anaeróbico. Na década seguinte, Wasserman e colaboradores consolidaram o termo “limiar anaeróbico”, definindo-o a partir de alterações ventilatórias observadas durante testes incrementais. Paralelamente, o grupo de fisiologia do esporte de Colônia, liderado por Alois Mader a partir de 1976, revolucionou a prática ao desenvolver métodos precisos e acessíveis de mensuração do lactato sanguíneo. Utilizando apenas 20 microlitros de sangue capilar, tornou-se possível monitorar alterações metabólicas em tempo real, transformando a análise de lactato em uma ferramenta rotineira de avaliação do desempenho.
Os pesquisadores de Colônia introduziram conceitos fundamentais para o treinamento esportivo, entre eles o uso da concentração de 4 mmol·L⁻¹ de lactato como estimativa do máximo estado estacionário de lactato (maxLass), e a classificação tripartida do treinamento em zonas abaixo, no e acima do limiar. Essas referências mostraram-se mais sensíveis às variações de performance de endurance do que parâmetros tradicionais como o V̇O₂max, permitindo maior precisão na prescrição de carga e intensidade.
Em 1984, Mader propôs um modelo abrangente de simulação do metabolismo energético humano, integrando as vias de resíntese de ATP — fosfocreatina, glicólise, fosforilação oxidativa e oxidação de lactato — com mecanismos autorregulatórios finamente ajustados. Nesse modelo, o ADP e o AMP atuam como ativadores de enzimas-chave, enquanto ATP e íons H⁺ exercem efeitos inibitórios, reproduzindo a dinâmica real observada durante o exercício. Essa abordagem permitiu descrever matematicamente as flutuações de ATP, ADP, AMP, fosfocreatina, lactato e pH, estabelecendo as bases de uma fisiologia quantitativa do esforço.
Uma das contribuições mais notáveis do modelo de Mader foi elucidar a base metabólica do maxLass e sua relação com a regulação da oxidação de gordura. Em intensidades inferiores ao maxLass, a produção glicolítica de piruvato é inferior à capacidade oxidativa mitocondrial, de modo que a demanda de acetil-CoA pode ser suprida predominantemente por ácidos graxos. No ponto de transição, a produção de piruvato iguala-se à sua taxa máxima de oxidação, caracterizando o equilíbrio entre os metabolismos aeróbico e anaeróbico. Acima desse ponto, o excesso de piruvato é convertido em lactato, provocando acidose metabólica, queda do pH e ativação do chamado slow component do V̇O₂, um aumento progressivo na ventilação e na demanda oxidativa para sustentar a resíntese de ATP.
Outro avanço conceitual decisivo foi a introdução da taxa máxima de glicólise ou formação de lactato (νLa.max), que expressa a capacidade de um atleta de gerar energia anaeróbica em curto prazo. Modelos computacionais demonstraram que indivíduos com νLa.max elevado atingem o maxLass em cargas menores, apresentando desempenho inferior em eventos de longa duração, mesmo quando possuem V̇O₂max similar a atletas de perfil mais aeróbico. Em contrapartida, esportes de curta duração e alta intensidade dependem de valores elevados de νLa.max para maximizar a potência. Assim, a interação entre maxLass e νLa.max tornou-se uma chave para interpretar o equilíbrio entre resistência e explosividade no contexto do treinamento.
A análise da deriva da curva de lactato representa outro refinamento conceitual importante. Essa curva, obtida em testes incrementais que relacionam intensidade de exercício à concentração de lactato sanguíneo, reflete o equilíbrio dinâmico entre produção e depuração de lactato. Alterações em sua forma — um deslocamento para a direita ou para a esquerda — indicam mudanças no estado metabólico do atleta, mas exigem interpretação cuidadosa. Um deslocamento para a direita, caracterizado por menor concentração de lactato para uma mesma intensidade, costuma refletir aumento da eficiência aeróbica, melhor economia de movimento ou adaptações corporais favoráveis. Já uma deriva para a esquerda pode ocorrer por destreinamento ou aumento da potência anaeróbica, sendo benéfica apenas em modalidades que dependem de esforços explosivos.
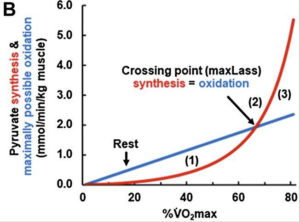
Legenda: (1) corresponde à zona em que a síntese glicolítica de piruvato está abaixo da taxa máxima possível de oxidação de piruvato pela fosforilação oxidativa. Nessa área, o déficit de piruvato permite que a oxidação de gorduras supra a demanda de substrato do metabolismo oxidativo ou que o piruvato seja obtido a partir do lactato previamente acumulado.
O ponto (2) é o ponto de intersecção em que a síntese glicolítica de piruvato e lactato iguala a taxa máxima possível de oxidação de piruvato para uma determinada carga de trabalho. Esse ponto indica o maxLass (máximo estado de equilíbrio de lactato). Em contraste, (3) representa a zona em que o fluxo glicolítico e a consequente síntese de piruvato excedem a taxa máxima possível de oxidação de piruvato. Como consequência, piruvato e lactato se acumulam. Extraído de Wackerhage H, Gehlert S, Schulz H, Weber S, Ring-Dimitriou S, Heine O. Lactate Thresholds and the Simulation of Human Energy Metabolism: Contributions by the Cologne Sports Medicine Group in the 1970s and 1980s. Front Physiol. 2022 Jul 22;13:899670.
O vínculo entre o metabolismo do lactato e as respostas ventilatórias, por sua vez, tornou-se um dos pilares da fisiologia aplicada. Os limiares ventilatórios, denominados VT1 e VT2, representam pontos específicos em que a ventilação aumenta desproporcionalmente em relação ao consumo de oxigênio (V̇O₂), refletindo alterações na produção e no acúmulo de lactato. O VT1, ou primeiro limiar ventilatório, marca o início do acúmulo mensurável de lactato no sangue, sinalizando a crescente participação do metabolismo anaeróbico. O VT2, ou segundo limiar ventilatório, corresponde à intensidade na qual a ventilação aumenta abruptamente, coincidindo com a perda do equilíbrio entre produção e remoção de lactato e aproximando-se do conceito de maxLass. Em média, o VT1 associa-se a concentrações em torno de 2 mmol·L⁻¹ e o VT2 a aproximadamente 4 mmol·L⁻¹, embora variações individuais sejam amplas. Essa correlação, baseada em mecanismos fisiológicos robustos, advém da ação dos íons H⁺ liberados durante a dissociação do ácido lático, que estimulam quimiorreceptores e intensificam a ventilação como forma de compensação da acidose metabólica.

Entretanto, essa associação não é isenta de críticas. A variabilidade individual na sensibilidade ventilatória, nas estratégias de tamponamento e na eficiência de remoção de lactato pode alterar substancialmente a coincidência entre limiares ventilatórios e metabólicos. Além disso, fatores externos como temperatura, altitude e desidratação influenciam a ventilação independentemente das mudanças metabólicas subjacentes. Também há limitações metodológicas nos protocolos de detecção dos limiares, o que contribui para discrepâncias entre estudos e aplicações práticas. Em esportes intermitentes, por exemplo, a ventilação pode não acompanhar com fidelidade as rápidas oscilações na produção de lactato.
Apesar dessas restrições, a análise integrada dos limiares ventilatórios e de lactato oferece vantagens notáveis. Ela permite a estimativa não invasiva das zonas de treinamento, facilita a individualização das cargas e contribui para o monitoramento da fadiga e da recuperação. Quando combinadas a medidas de frequência cardíaca e percepção subjetiva de esforço, as respostas ventilatórias e metabólicas compõem um retrato abrangente do estado fisiológico do atleta, fornecendo subsídios para decisões precisas sobre volume, intensidade e recuperação.
A contribuição do grupo de Colônia ultrapassa, portanto, o domínio teórico e permanece central na prática contemporânea do treinamento esportivo. Ao integrar fisiologia experimental, modelagem matemática e aplicação prática, Mader e seus colaboradores transformaram o lactato — antes visto apenas como subproduto metabólico — em uma janela privilegiada para observar o funcionamento energético do corpo humano. A análise da curva de lactato, da νLa.max, do maxLass e dos limiares ventilatórios continua sendo um dos métodos mais refinados para traduzir a complexa bioenergética do exercício em estratégias de treinamento eficazes. Essa herança científica consolidou as bases da fisiologia moderna do esporte e segue orientando a prescrição e o controle do treinamento com precisão, segurança e rigor fisiológico.
Referências:
Garcia-Tabar I, Gorostiaga EM. A “Blood Relationship” Between the Overlooked Minimum Lactate Equivalent and Maximal Lactate Steady State in Trained Runners. Back to the Old Days? Front Physiol. 2018 Jul 31;9:1034.
Wackerhage H, Gehlert S, Schulz H, Weber S, Ring-Dimitriou S, Heine O. Lactate Thresholds and the Simulation of Human Energy Metabolism: Contributions by the Cologne Sports Medicine Group in the 1970s and 1980s. Front Physiol. 2022 Jul 22;13:899670.
Wackerhage H. Contributions by the Cologne group to the development of lactate exercise testing and anaerobic threshold concepts in the 1970s and 1980s. J Physiol. 2021 Mar;599(5):1713-1714.
Wasserman K. The anaerobic threshold measurement to evaluate exercise performance. Am Rev Respir Dis. 1984 Feb;129(2 Pt 2):S35-40.
Os conceitos do limiar de lactato a serviço da evolução do atleta